


To characterize metabolites across the range of maternal glucose by comparing metabolomic profiles of mothers with high and low fasting plasma glucose (FPG).
We compared fasting serum from an oral glucose tolerance test at ∼28 weeks’ gestation from 67 Northern European ancestry mothers from the Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study with high (>90th percentile) FPG with 50 mothers with low (<10th percentile) FPG but comparable BMI. Metabolic data from biochemical analyses of conventional clinical metabolites, targeted mass spectrometry (MS)-based measurement of amino acids, and nontargeted gas chromatography/MS were subjected to per-metabolite analyses and collective pathway analyses using Unipathway annotation.
High-FPG mothers had a metabolic profile consistent with insulin resistance including higher triglycerides, 3-hydroxybutyrate, and amino acids including alanine, proline, and branched-chain amino acids (false discovery rate [FDR]-adjusted P < 0.05). Lower 1,5-anhydroglucitol in high-FPG mothers suggested recent hyperglycemic excursions (FDR-adjusted P < 0.05). Pathway analyses indicated differences in amino acid degradation pathways for the two groups (FDR-adjusted P < 0.05), consistent with population-based findings in nonpregnant populations. Exploratory analyses with newborn outcomes indicated positive associations for maternal triglycerides with neonatal sum of skinfolds and cord C-peptide and a negative association between maternal glycine and cord C-peptide (P < 0.05).
Metabolomics reveals perturbations in metabolism of major macronutrients and amino acid degradation pathways in high- versus low-FPG mothers.
Introduction
Maternal glucose metabolism during pregnancy differs from the pregravid state, as the mother must meet both her own and the growing fetus’s energy needs in a setting of increasing insulin resistance (1,2). Fasting glucose decreases progressively throughout gestation, but insulin resistance increases from the end of the first trimester through the third trimester. As insulin resistance increases, basal and stimulated insulin secretion, postprandial glucose levels, and hepatic glucose production all increase compared with the nongravid state. Maternal hyperglycemia occurs because of greater insulin resistance together with the inability of β-cells to meet the insulin secretory demands of increased insulin resistance (3). Although placental secretion of cytokines and other factors are known to contribute to pregnancy-induced insulin resistance (4), the mechanisms underlying the insulin resistance and its metabolic consequences in pregnancy remain incompletely defined.
Rapidly developing metabolomics technologies offer an integrative “omics” perspective into human physiology. Targeted assays of chemically similar groups of metabolites, e.g., amino acids, using stable isotope–labeled internal standards, afford precise measurement of specific metabolites. Complementary nontargeted gas chromatography (GC)/mass spectrometry (MS)–based assays provide more encompassing summaries by measuring features defined by mass spectra and retention time, with subsequent compound identification via comparison with a library of possible matches (5). Reflecting contributions of both genetic and environmental factors (5), the metabolome is uniquely relevant for defining metabolic changes characteristic of maternal hyperglycemia in an effort to better define pregnancy-induced insulin resistance and its consequences.
Given the importance of maternal glycemia on both the intrauterine milieu and long-term maternal outcomes, we sought to characterize the metabolic milieu associated with maternal hyperglycemia. We sampled 117 mothers from the Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study (6) with fasting plasma glucose (FPG) levels at the highest and lowest ends of the FPG distribution. Mothers with high FPG levels met criteria for gestational diabetes mellitus (GDM) using the new International Association of Diabetes and Pregnancy Study Groups (IADPSG) definition (7). Applied together, targeted and nontargeted metabolomics followed by pathway analyses point to metabolic cascades associated with maternal glycemia and shed light on factors underlying maternal insulin resistance, altered insulin secretion, and resulting hyperglycemia.
Research Design and Methods
We studied 117 HAPO mothers of northern European ancestry: 67 with high (>90th percentile) and 50 with low (<10th percentile) FPG levels according to the HAPO FPG distribution (6). Women were sampled to maintain comparable BMI, height, and gestational age for the two groups and were selected from the following HAPO field centers: Chicago, Illinois; Providence, Rhode Island; Cleveland, Ohio; Toronto, Canada; Belfast, U.K.; and Brisbane, Australia (Table 1).

Data and Sample Collection
HAPO study methods were described previously (6,8). Eligible women underwent a 75-g oral glucose tolerance test (OGTT) between 24 and 32 weeks’ gestation. Fasting, 1-h, and 2-h plasma glucose (PG); fasting and 1-h C-peptide; and HbA1c were measured. Additional fasting serum samples collected at the start of the OGTT were stored at −80° C until the present metabolomics assays.
Insulin sensitivity using OGTT glucose and C-peptide (ISOGTT C-pep) was calculated according to the methodology of Radaelli et al. (9), with a numerator adjustment for scaling: ISOGTT C-pep = 1,000/√(FPG × fasting C-peptide × G × C), where G and C are the means of fasting and 1-h PG (mg/dL) and C-peptide (μg/L), respectively.
Maternal height, maternal weight, newborn birth weight, newborn sum of skinfolds, and cord blood glucose and C-peptide were measured using standard procedures and calibrated equipment, and gestational age was determined as previously described (6,8). Demographic and lifestyle characteristics, age, parity, and self-identified ethnicity were collected via questionnaire.
Participants, caregivers, and HAPO study staff (except laboratory personnel) remained blinded to glucose values unless FPG was >5.8 mmol/L, 2-h OGTT PG was >11.1 mmol/L, random PG was ≥8.9 mmol/L, or any PG value was <2.5 mmol/L. Unblinded participants were excluded.
Conventional Metabolite Analyses
Conventional metabolites were measured on a Beckman-Coulter DxC600 autoanalyzer using reagents from Beckman (Brea, CA) (triglycerides and lactate) and Wako USA (Richmond, VA) (3-hydroxybutyrate). For free glycerol, we modified reagents by Roche (Indianapolis, IN) for glycerol-blanked triglycerides. To 84 mL of the Roche R1 reagent, we added 6.0 mg 4-aminoantipyrine dye (Sigma, St. Louis, MO). This assay was run by combining 250 μL reagent with 20 μL sample volume, calibrated against a glycerol standard (2.29 mmol/L) with detection at 520 nm after 5 min.
Targeted and Nontargeted Metabolomics Assays
Targeted assays of amino acids using stable isotope–labeled internal standards were performed on an Acquity TQD Triple Quadrupole system (Waters, Milford, MA) (10). Methanolic supernatants from the amino acid assay were spiked with a retention-time-lock (RTL) internal standard of perdeuterated myristic acid, dried, prepared for nontargeted GC/MS by methoximation and trimethylsilylation (11), and run on a 6890N GC-5975 Inert MS (Agilent Technologies, Santa Clara, CA). Programmed-temperature vaporization in the inlet and postrun, mid-column, hot backflushing of the GC column minimized analyte decomposition, carryover, and fouling of GC and MS. GC/MS data were deconvoluted with AMDIS freeware, courtesy of the National Institute of Standards and Technology, Gaithersburg, Maryland (12), and parsed against peaks annotated using the Fiehn RTL spectral library (11) with additions from our laboratory. Metabolites common across samples were tabulated, with peak areas log transformed and adjusted by scaling medians across two batches that differed due to temporal differences in sample analysis. Manual curation included correction of miscalls and identification of coeluting groups of isomeric metabolites (e.g., pentitols, hexitols).
Statistical Analyses
High- Versus Low-FPG Group Comparisons
All statistical analyses were performed in R (2.15.2). Categorical variable frequencies and continuous variable means were compared for high- and low-FPG mothers using Fisher exact tests and two-sample t tests, respectively. Women from U.S. HAPO field centers were grouped for analyses.
Conventional Metabolites and Targeted Amino Acids
Means and SDs were calculated for conventional metabolites and targeted amino acids for high- and low-FPG groups. One low-FPG mother was excluded from targeted analyses owing to outlying values for all analytes. Adjusted mean differences were estimated using linear regression with metabolite as the outcome, high- versus low-FPG status as the predictor of interest, and additional covariates for field center (referent, U.S.), parity (referent, first child), maternal and gestational age at OGTT, and fasting sample storage time. Additional adjustment for BMI and height at OGTT and drinking and/or smoking status was also examined. Exploratory linear regression models included newborn birth weight, newborn sum of skinfolds, and cord blood glucose and C-peptide as covariates, with and without adjustment for maternal FPG group.
Nontargeted Metabolites
When detectable, GC/MS nontargeted metabolite abundance is reported as log-2–transformed peak areas. GC/MS values were frequently undetectable because of absence from the sample or presence below detectability. Metabolites were included for statistical analysis if levels were detected for at least half of either FPG group. High- versus low-FPG group differences were identified using a mixture model to jointly consider detectable metabolite levels and frequency of undetectable levels. The mixture model can be viewed as a combination of a logistic model for metabolite detectability and a linear model for metabolite abundance, with an iterative computational procedure to estimate coefficients for both models at the same time (13). Likelihood ratio nested model χ2 tests were used to compare full models including high- versus low-FPG indicator variables in both the logistic and linear models with reduced models without the FPG variable. The linear model portion of the mixture model also included adjustment for the variables used in conventional metabolite and targeted amino acid linear regression analyses. Within FPG group, 1,000 bootstrap samples were used to estimate parameter SEs. When fewer than five samples yielded undetectable metabolite levels, the logistic regression component of the mixture model was eliminated, yielding results consistent with straightforward linear models.
Power Calculations
With 67 women in the high-FPG group and 50 in the low-FPG group, we had 80% power at a nominal two-sided 1% Type I error to detect unadjusted differences in group means of ∼0.64 SDs. The Type I error rate was chosen to approximate false discovery rate (FDR) adjustment at P < 0.05 for conventional metabolite and targeted amino acid analyses rather than strict family-wise error rate control. Nontargeted analyses were not formally powered.
Pathway Analyses
UniPathway Database
Pathway analyses used UniPathway annotation (http://www.grenoble.prabi.fr/obiwarehouse/unipathway) (14). UniPathway maps compounds to enzymatic reactions and then assembles nondivergent successions of reactions into linear subpathways, which are assembled into larger pathways and superpathways in a hierarchical organization.
GC/MS metabolites were mapped to UniPathway linear subpathways (ULSs) by translating to SMILES chemical structure representation using online software (15) and identifying matching groups using text comparison. Due to isomerism, matching groups were often comprised of multiple possible matches, particularly for sugars. Each possible match was considered for pathway analyses.
Pathway Enrichment Analyses
Pathway enrichment analyses used a self-contained gene set enrichment approach (16) using ULSs as compound “sets.” For each ULS, the median mixture model likelihood ratio test statistic for all constituent compounds was compared with the distribution of medians for the same metabolites after random reshuffling of sample covariate vectors 1,000 times. Reported P values represent the number of permutation medians as or more extreme than the observed median and were FDR adjusted (17).
Results
High- Versus Low-FPG Group Comparisons
Characteristics of high- and low-FPG groups are reported in Table 1. Mean FPG levels were statistically different by design; higher 1-h and 2-h PG were also observed in the high- vs. low-FPG group. Fasting and 1-h C-peptide levels were also higher in the high-FPG group (P < 0.01). BMI was similar at 28 weeks’ gestation but, consistent with glucose and C-peptide differences, insulin sensitivity was approximately three times higher in low- relative to high-FPG mothers (P < 0.01). HbA1c was also higher in the high-FPG group (P < 0.01). Field center representation differed between the two groups (P = 0.01), and there were borderline differences in parity, maternal age at OGTT, and sample storage time for high- versus low-FPG mothers. Rates of smoking or drinking, height, and gestational age at OGTT were comparable. Offspring of mothers in the high-FPG group had higher birth weight and cord C-peptide, although sum of skinfolds and cord glucose did not differ between the groups.
Conventional Metabolites and Targeted Amino Acids
For conventional metabolites, high-FPG mothers had higher levels of triglycerides and 3-hydroxybutyrate than low-FPG mothers (Table 2). Unadjusted differences remained statistically significant after covariate adjustment and FDR correction. Lactate and glycerol levels trended higher in the high-FPG group.
Conventional metabolite and targeted amino acid levels and adjusted differences in HAPO mothers with high and low FPG
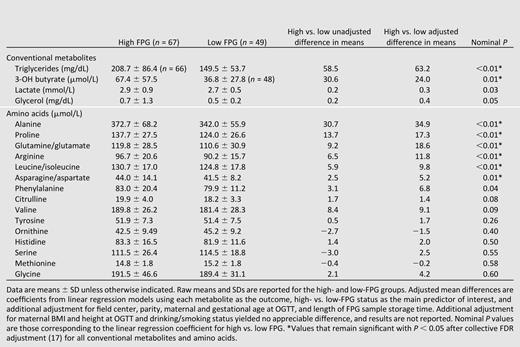
Among targeted amino acids, alanine, proline, glutamine/glutamate, arginine, leucine/isoleucine, and asparagine/aspartate levels were higher in high- versus low-FPG mothers, with differences remaining statistically significant after covariate adjustment and FDR correction (Table 2). Phenylalanine, citrulline, and valine trended higher in high- versus low-FPG mothers. The remaining targeted amino acids were not significantly different in high- versus low-FPG groups.
Nontargeted GC/MS Metabolites
Of 309 compounds identified in nontargeted assays, 130 were detectable in at least half of the samples from either FPG group. The mixture model identified 30 compounds/classes differing in detectability or abundance in the high- versus low-FPG groups, with FDR-corrected P < 0.05 (Table 3).
Nontargeted metabolite estimated proportions of present metabolites, estimated mean detected levels, and adjusted differences in HAPO mothers with high and low FPG
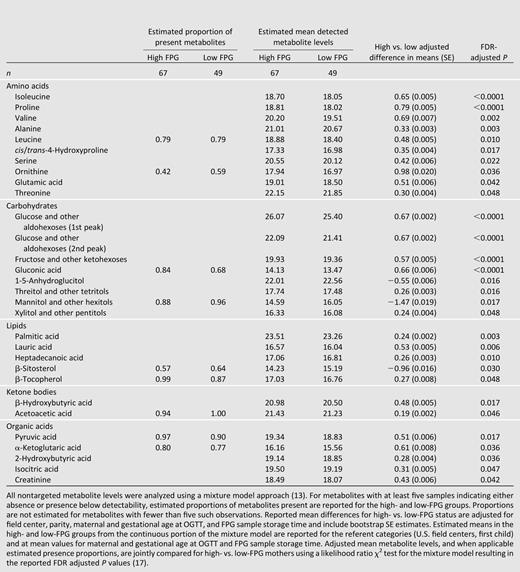
Nontargeted GC/MS results confirmed and augmented observations made with the targeted platform. Higher proline and isoleucine levels were observed in high-FPG mothers in both platforms. The trend toward a higher valine level in the high-FPG group observed for targeted data was confirmed by nontargeted data. While most nontargeted metabolites with statistically significant differences were higher in high-FPG mothers, 1,5-anhydroglucitol was among the few with lower levels for high- vs. low-FPG mothers.
Pathways
To better define metabolic perturbations in high vs. low FPG mothers, pathway analyses were performed to complement per-metabolite analyses by identifying metabolic pathways that exhibited collective differences in metabolite levels between groups. Collective differences in high- versus low-FPG mothers were observed for 14 unique ULSs (FDR-adjusted P < 0.05 [Supplementary Table 1]). The majority of these ULSs are involved with amino acid metabolism and feed into pathways contributing to proline biosynthesis and degradation as well as leucine, phenylalanine, lysine, and alanine degradation (Supplementary Figure 1). The other ULSs identified in Supplementary Table 1 contribute to carbohydrate and organic acid metabolism.
Newborn Associations
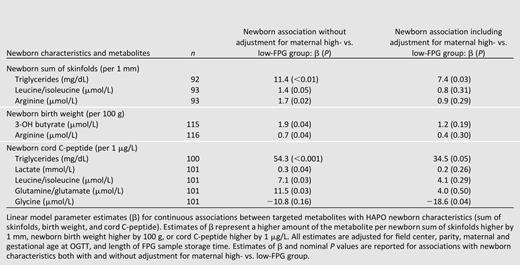
Exploratory analyses of maternal metabolite levels determined in the targeted assays and newborn phenotypes suggest positive associations for triglycerides, leucine/isoleucine, and arginine with sum of skinfolds, 3-hydroxybutyrate, and arginine with birth weight, as well as triglycerides, lactate, leucine/isoleucine, and glutamate with cord C-peptide (P < 0.05, Table 4). Owing to newborn outcome associations with maternal FPG, adjustment for maternal FPG group attenuated these observations, with only the association of maternal triglycerides and newborn sum of skinfolds and cord C-peptide retaining statistical significance (triglycerides higher by 7.4 and 34.5 mg/dL associated with sum of skinfolds higher by 1 mm, P < 0.03, and cord C-peptide higher by 1 μg/L, P < 0.05, respectively). Interestingly, after adjustment for maternal FPG group, maternal glycine levels demonstrated a negative association with cord C-peptide (Table 4). No associations between maternal metabolite levels and cord glucose were observed with P < 0.05 with or without adjustment for maternal FPG group. Multiple metabolites in the nontargeted assays demonstrated association with sum of skinfolds or cord C-peptide, but all of these associations were attenuated after adjustment for maternal FPG group (Supplementary Table 2).
Conclusions
Broad-scale characterization of metabolic differences associated with hyperglycemia and insulin resistance during pregnancy is possible given recent developments in metabolomics technology. Two recent metabolomics studies, one using nuclear magnetic resonance spectroscopy and the other liquid chromatography–MS (LC/MS), examined urine from women with GDM compared with healthy pregnant control subjects. In one, GDM women with pronounced hyperglycemia had higher levels of glucose, lysine, and citrate, while a second smaller study found higher levels of choline and a metabolite of leucine (18,19). However, the latter findings were not confirmed in a different study using LC/MS (20). We used targeted and nontargeted GC-MS–based methodologies to identify key metabolites in serum and metabolic pathways associated with hyperglycemia and insulin resistance at ∼28 weeks’ gestation to gain insights into factors contributing to maternal hyperglycemia. This was done by comparing mothers at the upper and lower ends of the distribution of FPG and insulin sensitivity. The high-FPG group included pregnant mothers who meet recently adopted IADPSG criteria for GDM, suggesting impaired β-cell function together with insulin resistance (7). To our knowledge, this is the first report using targeted and nontargeted metabolic profiling to characterize serum from hyperglycemic, insulin-resistant mothers during pregnancy and more insulin-sensitive, low-FPG counterparts.
Analyses of conventional metabolites in high- vs. low-FPG mothers confirmed expected differences associated with differences in insulin sensitivity, specifically higher triglyceride and 3-hydroxybutyrate levels, with a similar trend for lactate and glycerol. Pregnancy is characterized by an increase in triglycerides compared with the nongravid state, with higher levels in GDM mothers (1). The finding of higher triglyceride levels in the high-FPG group likely reflects, in part, greater insulin resistance and probable impaired insulin secretion in this group. Insulin resistance may have also contributed to the higher lactate and 3-hydroxybutyrate levels, as a recent population-based study in nonpregnant individuals reported association of these metabolites with insulin sensitivity (21). The higher level of lactate in the high-FPG group is also consistent with its role as a substrate for gluconeogenesis.
Pregnancy is also characterized by hypoaminoacidemia with marked decreases in gluconeogenic amino acids with fasting during pregnancy (2). Previous studies have demonstrated differences in levels of specific amino acids between GDM and non-GDM mothers (22–24). Consistent with findings in some but not all studies of women with GDM (23,24), we found higher alanine levels in high- versus low-FPG mothers, consistent with alanine being a major substrate for gluconeogenesis in the setting of maternal hyperglycemia. Higher levels of proline, glutamine/glutamate, and asparagine/aspartate were also present in the high-FPG mothers, consistent with recent population-based and other studies in nonpregnant cohorts that demonstrated association of higher levels of these amino acids with glucose levels, insulin resistance, or obesity (21,25,26).
An area of increasing interest is the role of branched-chain amino acids (BCAAs) in insulin resistance, hyperglycemia, and type 2 diabetes—an association first noted >30 years ago (25,27). More recent cohort and/or population-based studies have demonstrated association of BCAA levels with insulin resistance, higher glucose levels, and changes in insulin resistance and glucose homeostasis after weight loss (21,25,26,28,29), while higher levels of BCAAs are present in type 2 diabetic subjects compared with control subjects (30,31) and are part of a metabolic signature that predicts incident type 2 diabetes (26,32,33). Some, but not all, prior studies have demonstrated higher levels of BCAAs in GDM (22–24). In the current study, the targeted and nontargeted analyses demonstrated higher levels of BCAA in high-FPG mothers, consistent with findings in nonpregnant populations and suggestive of similarities between hyperglycemia and insulin resistance and its consequences in pregnant and nonpregnant populations.
Pathway analyses using nontargeted data also suggested differences in amino acid metabolism between the high- and low-FPG groups, including altered degradation of proline, phenylalanine, lysine, leucine, and alanine. Taken together, the data in the current study have now extended previously reported associations in nonpregnant populations by demonstrating that, during pregnancy, greater insulin resistance together with hyperglycemia, reflecting accompanying impaired insulin secretion, is associated with levels of multiple amino acids and altered activity of amino acid degradation pathways. As amino acids are actively transported from the mother across the placenta and placental amino acid exchange may be altered in GDM (2,23), alterations in placental transport may also contribute to differences in amino acid levels between the high- and low-FPG group.
Previous metabolomics studies of insulin resistance focused largely on obese, insulin-resistant compared with lean, insulin-sensitive individuals (25,28,29). The novelty of the current study is that, while BMI was similar between high- and low-FPG groups, the high-FPG group with greater pregnancy-induced insulin resistance exhibited evidence for amino acid differences similar to those observed in obese, insulin-resistant individuals. The persistence of these amino acid differences suggests they are characteristic of insulin resistance independent of obesity. As the hyperglycemic mothers also met new criteria for GDM, some degree of β-cell dysfunction and impaired insulin secretion also may have contributed to observed differences between groups. In HAPO, maternal obesity and hyperglycemia appeared to have independent effects on newborn outcomes (34). Determining whether maternal obesity in the absence of hyperglycemia leads to changes in the metabolic profile similar to or different than those associated with high glucose levels and insulin resistance will require future studies.
Another novel aspect of the current study is inclusion of nontargeted metabolomics profiling of high- versus low-FPG mothers. As expected based on calculated insulin sensitivity in the high- versus low-FPG groups, nontargeted GC/MS data together with targeted analyses demonstrated a metabolic profile consistent with insulin resistance and, possibly, impaired insulin secretion in the high-FPG group. This included higher levels of gluconate (perhaps reflecting hyperglycemic excursions and increased oxidative stress), free fatty acids (indicative of impaired suppression of lipolysis by insulin), by-products of fatty acid oxidation (ketones and tricarboxylic acid cycle intermediates), multiple amino acids, and fructose, pentitols, and hexitols. Simultaneous elevation in triglycerides, gluconeogenic precursors, and assorted sugars is consistent with a recent description of insulin resistance “selective” for failure to suppress gluconeogenesis but with normal anabolic effects of insulin on lipogenesis in liver (35).
Among the few nontargeted analytes with significantly lower levels in high- versus low-FPG mothers in the current study was 1,5-anhydroglucitol. Fluctuation of 1,5-anhydroglucitol in response to hyperglycemic excursions has been reported, with lower levels in diabetic subjects with poor glycemic control, ostensibly because glucose competes with 1,5-anhydroglucitol for renal uptake (36). Among pregnant women with type 1 diabetes, 1,5-anhydroglucitol levels are highly correlated with glucose levels determined by continuous glucose monitoring and, in the third trimester, the best predictor of macrosomia (37). The nontargeted assays suggest that 1,5-anhydroglucitol may also have clinical utility as a marker of maternal hyperglycemia in pregnancy.
The current study was not specifically designed to examine the impact of maternal metabolites on newborn outcomes, but exploratory analyses were conducted. Arginine, leucine/isoleucine, glutamine, 3-hydroxybutyrate, lactate, and triglycerides demonstrated trends toward association with cord C-peptide, birth weight, or sum of skinfolds (a measure of newborn adiposity) in targeted assays. In the nontargeted assays, a variety of sugars and their degradation products also demonstrated trends toward association with sum of skinfolds or cord C-peptide levels. Only association of maternal triglycerides with cord C-peptide levels and neonatal sum of skinfolds remained significant after adjustment for maternal FPG group, although in the current study sum of skinfolds did not differ in the offspring despite the higher birth weight in offspring of the high-FPG group. These data are consistent with the suggested role of triglycerides on fetal size at birth in the settings of normal glucose tolerance and GDM (38,39). Interestingly, maternal glycine demonstrated negative association with cord C-peptide after adjustment for maternal FPG group, consistent with glycine’s negative association with insulin resistance in nonpregnant populations (40).
This study had numerous strengths. The HAPO Study was population based, and the large number of participants allowed for select groups to be identified for study. Moreover, HAPO used a common protocol and data collection instruments, uniform training of personnel, and a central laboratory for analyses to ensure consistency of results. One limitation of a large study is that limited measures were obtainable; e.g., measures of maternal body fat and its distribution are not available. An additional limitation is that findings of nontargeted assays require development of specific targeted assays for metabolites of interest to confirm findings. Finally, the current study examined women from the upper and lower ends of the distribution of FPG. Future studies will need to examine women across the continuum of glucose levels to confirm the reported findings and more fully characterize the maternal metabolome and its impact on newborn outcomes.
The mechanisms underlying maternal insulin resistance and its metabolic consequences remain incompletely defined. The current study provides new data about the metabolic profile characteristic of maternal hyperglycemia and suggests a role for amino acids in insulin resistance and hyperglycemia during pregnancy, similar to nonpregnant populations. As the current study is an initial investigation comparing upper and lower ends of the FPG and insulin sensitivity distribution, differences between the two groups may reflect not only insulin resistance and β-cell dysfunction in the high-FPG group but also potentially a lesser degree of pregnancy-induced insulin resistance in the low-FPG group. Future population-based studies examining pregnant women across the full continuum of glucose levels, insulin sensitivity, and BMI will provide opportunity to more fully characterize metabolic profiles associated with maternal hyperglycemia, insulin resistance, and obesity and their association with newborn outcomes. This initial investigation demonstrates broad-scale metabolic differences associated with hyperglycemia and insulin resistance in pregnancy that help to provide insight into the underlying mechanisms and metabolic consequences of pregnancy-induced insulin resistance and identify potential new markers of maternal hyperglycemia.
Acknowledgments.
The authors thank Stephan Baumann and Dr. Steven Fischer at Agilent Technologies, Inc., for assistance in developing the nontargeted GC/MS platform.
Funding. This study was funded by grants R01-HD34242 and R01-HD34243 from the National Institute of Child Health and Human Development and the National Institute of Diabetes and Digestive and Kidney Diseases, by the National Center for Research Resources (M01-RR00048, M01-RR00080), by the National Institute on Aging (P30-AG028716), and by the American Diabetes Association and Friends of Prentice.
Duality of Interest. No potential conflicts of interest relevant to this article were reported.
Author Contributions. D.M.S. contributed to the study design, data analysis, and interpretation of findings and primary drafting of the manuscript. M.J.M. was responsible for assays of conventional metabolites and nontargeted metabolomics, interpretation of findings, and manuscript writing. N.R.D. contributed to data analysis. R.D.S. developed and performed targeted metabolomics assays. A.R.D. contributed to study design and statistical modeling. L.P.L. contributed to study design. B.E.M. contributed to study design and interpretation of findings. C.B.N. contributed to study design, interpretation of findings, and manuscript writing. J.R.B. was responsible for assays of conventional metabolites and nontargeted metabolomics, interpretation of findings, and manuscript writing. W.L.L. conceived the hypothesis and contributed to study design, interpretation of findings, and manuscript writing. D.M.S. and W.L.L. are the guarantors of this work and, as such, had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis.
Prior Presentation. Parts of this study were presented in abstract form at the 72nd Scientific Sessions of the American Diabetes Association, Philadelphia, Pennsylvania, 8–12 June 2012.
